"人体中的导航仪"——大脑的空间表征机制简介
北京的初冬是个美丽的时节,你决定去动物园散散心。可是和很多人一样,你是一个不折不扣的"路痴":"找不着北",没有方向感;虽然已经去过动物园几次了,却还是找不到位置;并且,随着离学校越来越远,你发现你已经找不到回去的路,迷失在了茫茫人海之中。
最终,无奈的你决定叫一辆出租车。坐在出租车上,你看到了刚才走过的一些道路,还有一些是你上次来时见到的,在一些熟悉的路口准确的转向后,你终于来到了动物园。这时,你不禁由衷地钦佩出租车司机,觉得拥有良好的空间感真是一件了不起的事。可是,为什么出租车司机能够找到准确的位置呢?他在上海还能找的这样准吗?这种能力的生理机制是什么?要想解决这些问题,我们必须要从认识大脑中的空间表征机制开始。
从上面的经历可以看到,要想准确地到达目的地并不是一件简单的事,其中涉及到了复杂的认知过程,包括对视、听觉信息的加工处理,对场景的学习和对空间记忆的提取等等。从生理角度来讲,也就是需要各个脑区的协同运作,共同实现定位和导向等任务。既然如此,我们的神经系统中是否存在专门负责空间表征的区域呢?这是科学家们首先想解决的问题。
1948年,实验心理学家托尔曼发现,大鼠在迷宫中达到特定目标的过程,表现得比单纯的感觉运动反应更为迅速直接,这说明大鼠脑内很有可能存在着一套对环境空间的认知机制,托尔曼把这种内生于脑内的心理表征称为"空间认知地图",他认为这一机制是大鼠和其他哺乳动物构建对环境的空间概念以及定位导向的主要手段(Tolman,1948)。
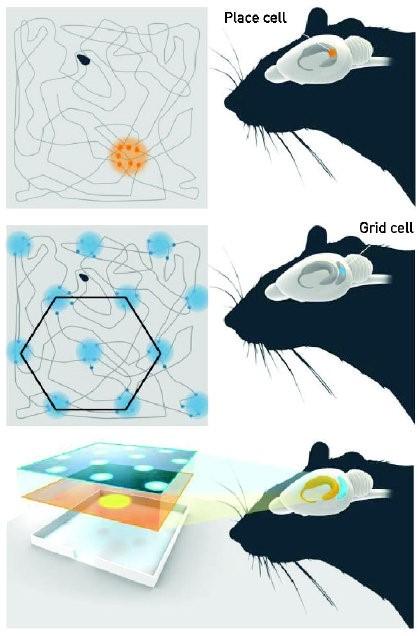
他的猜想随后被O'Keefe和Dostrovsky证实(O'Keefe & Dostrovsky, 1971)。他们通过对大鼠的单神经元电生理记录发现,在海马体的CA1区存在着一种对特定空间位置兴奋的细胞,O'Keefe等人称之为位置细胞(place cell)。在给定的空间场景中,位置细胞会对应一块特定的位置野(place field),当大鼠位于这一位置上时,对应的位置细胞就会兴奋。当大鼠移动到不同位置,就会有不同的位置细胞兴奋。这样一来,场景中的位置信息就通过海马中的位置细胞映射到大鼠脑中,帮助大鼠形成了对于该场景下空间方位的认知。
O'Keefe等人随后又发现(O'Keefe, Burgess, Donnett, Jeffery & Maguire, 1998),不同位置细胞的相对位置关系并不对应于其位置野在真实空间中的位置关系,也就是说,两块相邻的位置野并不一定对应两个相邻的位置细胞,大鼠是通过整合不同位置细胞的兴奋信息实现对当下空间位置的整体认知的。在特定情境空间中,这样的位置细胞群可以通过学习形成,并较为稳定地保持一段时间,形成对空间的记忆。在不同空间情境中,感受兴奋的位置细胞群则是动态变化的,这样大鼠就能在脑中建立起许多不同的"认知地图",适应不断变化的空间场景。
位置细胞的发现为托尔曼的理论提供了强有力的证据,科学家们一度认为海马就是"空间认知地图"的生理结构所在。但位置细胞的功能局限并不支持这一观点,因为每一个位置细胞可以对应多个位置野,位置野交互重叠,无法精准反映不断变化的空间位置关系(O'Keffe & Burgess, 2005)。据Darwin于1873年提出的路径整合(path integration,PI)理论,动物在运动过程中,需要时刻保持相对于环境位置的矢量关系,不断更新的矢量关系为动物提供方位,间距等重要的空间信息。显然,单凭位置细胞无法精准地做到这一点。长期以来,研究者对空间整合机制知之甚少。
直到2005年,Hafting等人通过大鼠的实验箱觅食实验,在其背侧内嗅皮层(dorsal medial entorhinal cortex, dMEC)中发现了对特定空间位置强烈放电的网格细胞(grid cell),(Hafting, Fyhn, Molden, Moser & Moser, 2005),为路径整合机制的研究开启了新大门。网格细胞与位置细胞类似,都会在特定的位置野产生最大放电,但不同的是,网格细胞与位置野是一一对应的,通过不断加入新的网格细胞,位置野网络理论上能够无限延展。并且这种位置野网络分布更加有序,在增大实验环境,去除噪声后,研究者发现网格细胞的位置野分布近似于正六边形网格。这样的六边形网络在空间中相当稳定,实验者在旋转实验场地后发现,该网络在空间中也发生了等角度旋转。在黑暗条件下,虽然失去了外部线索,该网络的几何特点,放电频率和信息加工仍然能够保持稳定。网格细胞的这些特点有力支持了路径整合功能,像一幅坐标图,在背侧内嗅皮层和空间环境间建立了稳定的对应关系。
除了位置细胞和网格细胞,研究者还发现了一些其他与空间表征相关的细胞,包括Muller等人在大鼠头下托(presubiculum)发现的头向细胞(head direction cell),这种神经元能够在大脑朝向特定方向时产生最大放电(Muller, Ranck & Taube, 1996);O'Keefe等人在大脑多处发现的边界细胞(boundary cell),可以感知环境的边缘轮廓(O'Keefe & Burgess, 2006;O'Keefe & Burgess, 2009)。这些细胞被认为与位置细胞,网格细胞一起参与到了空间表征的过程。
目前一般认为,海马-嗅皮层回路是脑内空间定位导航系统的主要组成部分,但两者之间交互作用的机制,以及在空间表征中发挥作用的多个脑区间的相互关联机制目前还不甚清楚。已有的研究认为内嗅皮层的空间信息和嗅周皮层的非空间信息在海马内整合(Hargreaves, Rao, Lee & Knierim, 2005),当动物再次进入该环境时,储存在海马内的空间、非空间信息激活内嗅皮层的路径整合器(Witter & Moser, 2006),网格细胞通过自身及从运动中从头向细胞,边界细胞得到的速度、方向等信息获得实时场景位置的空间表征及自身的相对位置信息,并通过海马传入的信息进行及时反馈,调节网格细胞自身信息的累积错误以维持稳定的定位导向功能(Burgess, Barry & O'Keefe, 2007;Fyhn, Hafting, Treves, Moser & Moser, 2007;Kropff, Carmichael, Moser & Moser, 2015)。但这一机制的细节,还有待科学家们进一步的研究。
空间定位导向细胞的发现是科学的重大进步。这种不依赖于外界感觉输入的内生空间网络印证了哲学家Kant不依赖于经验的先验论,促进了我们在神经层面对动物空间认知和空间记忆功能的认识,在应用层面,由于早期阿兹海默氏病(AD)患者常伴随海马和内嗅皮层的功能损伤,这一发现也有助于我们更好地认识这一疾病的致病机理。
2014年诺贝尔生理学奖授予了对"发现与大脑定位系统相关的细胞"做出重大贡献的O'keefe以及Moser夫妇,但关于空间表征认知机制的研究还在继续。有人指出,人的空间方位识别、导向能力比老鼠要差得多,对人类来说,海马-嗅皮层环路结构的作用是十分有限的,那么,究竟是不是如此呢?是什么导致了你和出租车司机之间的差距,这些还有待我们更深入的研究。
参考文献
Moser, E. I., Kropff, E., & Moser, M. B. (2008). Place cells, grid cells, and the brain's spatial representation system. Annual Review of Neuroscience, 31(1), 69.
Kropff, E., Carmichael, J. E., Moser, M. B., & Moser, E. I. (2015). Speed cells in the medial entorhinal cortex. Nature, 523(7561), 419.
Tolman, E. C. (1948). Cognitive maps in rats and men. Psychological Review, 55(4), 189.
O'Keefe, J., & Dostrovsky, J. (1971). The hippocampus as a spatial map. preliminary evidence from unit activity in the freely-moving rat. Brain Research, 34(1), 171-175.
O'Keefe, J., Burgess, N., Donnett, J. G., Jeffery, K. J., & Maguire, E. A. (1998). Place cells, navigational accuracy, and the human hippocampus. Philosophical Transactions of the Royal Society of London, 353(1373), 1333.
Burgess, N., & O'Keefe, J. (2005). The theta rhythm. Hippocampus, 15(7), 825-826.
Hafting, T., Fyhn, M., Molden, S., Moser, M. B., & Moser, E. I. (2005). Microstructure of a spatial map in the entorhinal cortex. Nature, 436(7052), 801.
Muller, R. U., Jr, J. B. R., & Taube, J. S. (1996). Head direction cells: properties and functional significance. Current Opinion in Neurobiology, 6(2), 196-206.
Barry, C., Lever, C., Hayman, R., Hartley, T., Burton, S., & O'Keefe, J., et al. (2006). The boundary vector cell model of place cell firing and spatial memory. Reviews in the Neurosciences, 17(1-2), 71.
Lever, C., Burton, S., Jeewajee, A., O'Keefe, J., & Burgess, N. (2009). Boundary vector cells in the subiculum of the hippocampal formation. Journal of Neuroscience the Official Journal of the Society for Neuroscience, 29(31), 9771.
Hargreaves, E. L., Rao, G., Lee, I., & Knierim, J. J. (2005). Major dissociation between medial and lateral entorhinal input to dorsal hippocampus. Science, 308(5729), 1792.
Sargolini, F., Fyhn, M., Hafting, T., Mcnaughton, B. L., Witter, M. P., & Moser, M. B., et al. (2006). Conjunctive representation of position, direction, and velocity in entorhinal cortex. Science, 312(5774), 758.
Burgess, N., Barry, C., & O'Keefe, J. (2007). An oscillatory interference model of grid cell firing. Hippocampus, 17(9), 801.
Fyhn, M., Hafting, T., Treves, A., Moser, M. B., & Moser, E. I. (2007). Hippocampal remapping and grid realignment in entorhinal cortex. Nature, 446(7132), 190-4.
来源:知乎 www.zhihu.com
作者:Alexey Yoo
【知乎日报】千万用户的选择,做朋友圈里的新鲜事分享大牛。 点击下载
没有评论:
发表评论