
(找不到封面配图了,请大家赏花_(:з」∠)_。新鲜的,周末才拍出来的。)
1992~2002年间发表在Science和Nature上的一系列研究向人们揭示了CaMKII蛋白在学习和记忆中起到的作用。下面将回顾其中的几篇文章。
在1992年之前,研究者们就猜想,LTP可能是一种学习机制。当时支持这种猜想的证据只有一条,即阻断NMDA受体会导致空间学习能力受损。但是这条证据是有问题的,因为阻断NMDA受体会导致突触功能缺陷,从而干扰海马回路的功能。为了解决这个问题,Sliva等人想要在保持突触功能正常的情况下减少对突触激活水平,从而检验是否能消除LTP。而αCaMKII是一个被发现只存在于神经系统中的蛋白激酶,且它一旦自我磷酸化后就不再依赖于Ca+/CaM;一些学习模型中也发现了这种机制。也就是说,αCaMKII可能与LTP有关。
提一句,1992年7月的Science杂志被Sliva的组霸刊了,连发了两篇文章。一篇讲基因变异模型及细胞水平上进行的模型检验,一篇专门讲行为学水平上的模型检验。鉴于后者比较容易理解, 这里主要聚焦于第一篇文章。
老师说,现在想要发Science,可没那么容易了,醒醒吧别睡了。
1)αCaMKII变异模型
研究者采用基因打靶(gene target)技术在αCaMKII基因序列上的调节域中敲入一个抗新霉素的 neo 基因。这样一来,αCaMKII基因就无法正常表达出蛋白质了。
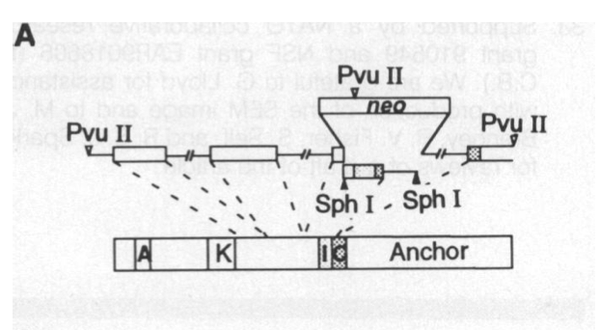
打靶载体通过电转移法 (electroporation) 被导入囊胚的胚胎干细胞。由于打靶载体带有抗新霉素的 neo 基因,整合了打靶载体的胚胎干细胞能够在有新霉素(neomycin)的环境中生长,继而被筛选出来。之后,这些干细胞打回囊胚中。
为了检验是否完全干扰了αCaMKII基因、阻断了其转译和表达,研究者先后做了南方检验(Southern analysis/blot)、西方检验(Western analysis)和北方检验(Northern analysis)。南方检验是对DNA的检验。
南方检验是对DNA的检验。上图是对野生型、变异杂合子和变异纯合子的DNA检验。Kb表示的是碱基对的多少。从图中可以看到,野生型只有单一碱基对长度的链;变异杂合子有一条和野生型一样长的链,还有两条短链;变异纯合子只有两条短链。这是因为变异链上新打入的neo基因带有限制性内切酶Pvu II的一个剪切位点,所以Pvu II可以在变异链的中间多咔嚓一刀,把一条长链变成两条短链。颜色的深浅与检验样本的质量大小有关。
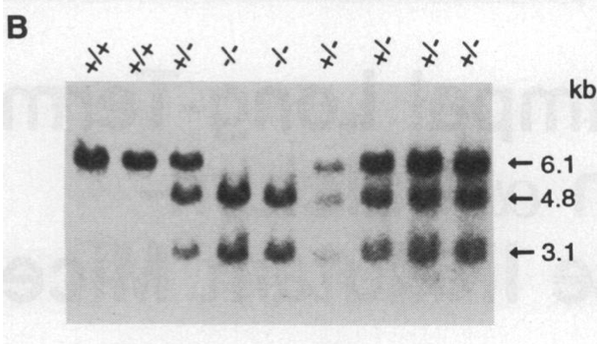
北方检验是对RNA进行分析。既然已经没有原表达基因了,DNA自然无法转译,为什么还要进行RNA分析?研究者是为了确认没有DNA残链被转译成了RNA(有可能残链转译的RNA也会被表达成功能性蛋白)。在当时(1992年),人们还不知道在原表达基因已经被破坏了的情况下,是不会有RNA被转译出来的。检验的结果表明,变异的样本中没有αCaMKII或其残缺版的mRNA,但是β-CaMKII的mRNA水平与正常小鼠的无异。
最后,西方检验是对蛋白质的检验。图中最右一栏的是对照蛋白样本。从图中可以看到,变异纯合子无αCaMKII,但有β-CaMKII。(注:beta-CaMKII有两种亚型,因此图中共有三个指示箭头;kD为蛋白质质量单位。)

至此,我们可以下结论说,αCaMKII的基因变异模型被建立起来了。接下来要对其进行功能性检验。
2)解剖学及基本行为表现检验
如果对野生型和变异型小鼠的海马区所在的前脑组织进行切片染色,你有什么预期?两者的海马区会有差别吗?有什么样的差别?
行为学上呢?外观、体重会有差别吗?基本的嗅闻行为(sniffing)、求偶行为(mating)和运动机能上会有差别吗?
当然啦,你大可以做出两者相差巨大的预期,毕竟发生了基因变异,其他基因的表达是否会受到影响谁也说不好。但是偏向着"成功模型"的预期是,两者在解剖学和基本的行为表现上不应该存在显著差异。不然要如何解释后面学习差异的原因呢?
幸运的是,确实没有多大的差别。变异小鼠(纯合子)的新皮层和海马皮层的细胞排列很正常,它们也会跑会跳会交配,没有运动失调症状(ataxia)。有显著性差别的地方仅在于两处,其中一处是变异小鼠似乎更神经质一些,对于人类抚摸更加回避。
另一处则是它们在用于衡量空间记忆能力的莫里斯水迷宫(Morris Maze)等实验任务中表现出明显的缺陷 。
3)突触功能检验:全细胞记录
为什么要进行突触功能的检验呢?之前说了嘛,就是因为以前的模型阻断了NMDA受体,会影响到突触功能,说不定对海马回路都产生了影响,所以无法说究竟什么才影响了记忆(当然了,NMDA也影响记忆,但还不是核心机制)。
这样一来,成功的预期也很明了了:变异的小鼠其突触功能应当完好。
结果当然是如你所料啦(毕竟文章也发出来了_(:з」∠)_):与野生小鼠相比,变异小鼠的突触后电位形状和峰值、阻断NMDA或非NMDA受体通道后的突触后电位峰值、电压依赖性曲线都没啥区别。也就是说,NMDA受体功能性在两者身上无差。
有区别的地方也是对模型有利的。在野生小鼠的脑片中(in vitro),如果给予一次刺激后紧接着发放第二次刺激,那么第二次刺激所诱发的突触后兴奋电位应当升高。这个现象称Paired-pulse facilitaion(成对刺激促进作用)。变异小鼠的脑切片也体现出了这种促进作用,但是明显不及野生型小鼠。也不奇怪,毕竟促进前膜突触囊泡释放的synapsin I分子,是由CaMKII调控的。
4)长时程增强作用(long-termpotentiation,LTP)受损
终于到了激动人心的时刻。
我们的预期也很简单了,变异小鼠的LTP大概是受损严重,甚至根本就消失了。毕竟它们在海马功能检验的一系列行为学测试中的表现令人失望(又令人满意)。
首先进行的是场电位记录(field potential recording),其记录的是许多神经元的活动,衡量的是突触强度(Synaptic strenghth)。突触强度,指的是给予强直性刺激后,突触后兴奋电位随时间的变化,用其与基线电位水平的百分比值表示。强直性刺激条件是100hz的20次刺激,每次刺激间隔11秒,共进行3个刺激串。结果如下图。
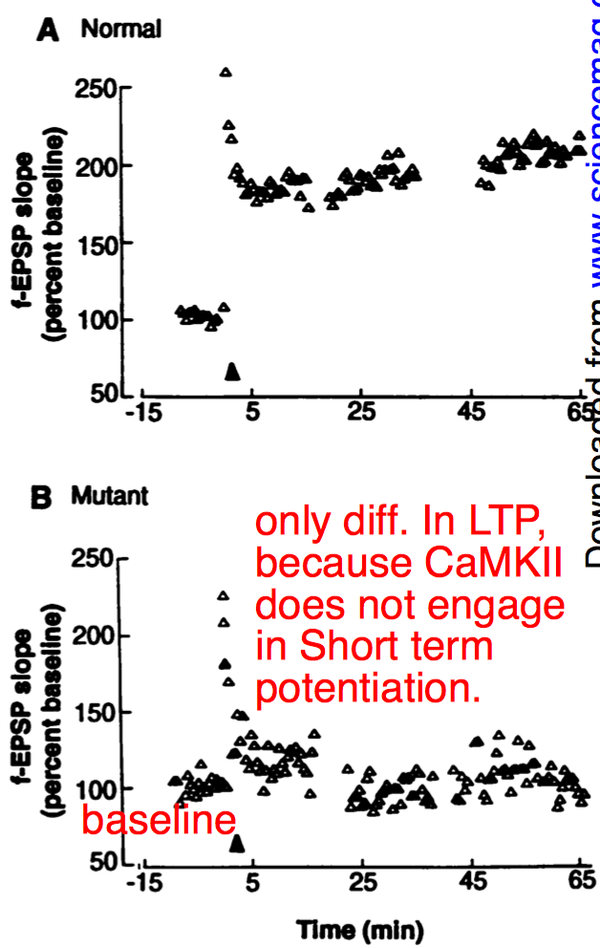
以给予刺激的时间点为时间轴零点。零点前记录的为基线水平。给予刺激后,场电位峰值无明显差异,但在接下来一个小时内的电位水平存在明显差异:野生型小鼠出现了长时程增强作用,而变异小鼠则很快回到基线水平。这很好理解,因为CaMKII不参与短时程增强作用(与突触前膜所释放的神经递质多少有关)。
接下来研究者对单个细胞进行了全细胞记录(whole cell recording)。结果和场电位记录是相似的。野生型小鼠在接受强直刺激后出现LTP,而变异小鼠则没有LTP。有意思的是,有两只变异纯合小鼠出现了LTP,但幅度明显不如野生型。
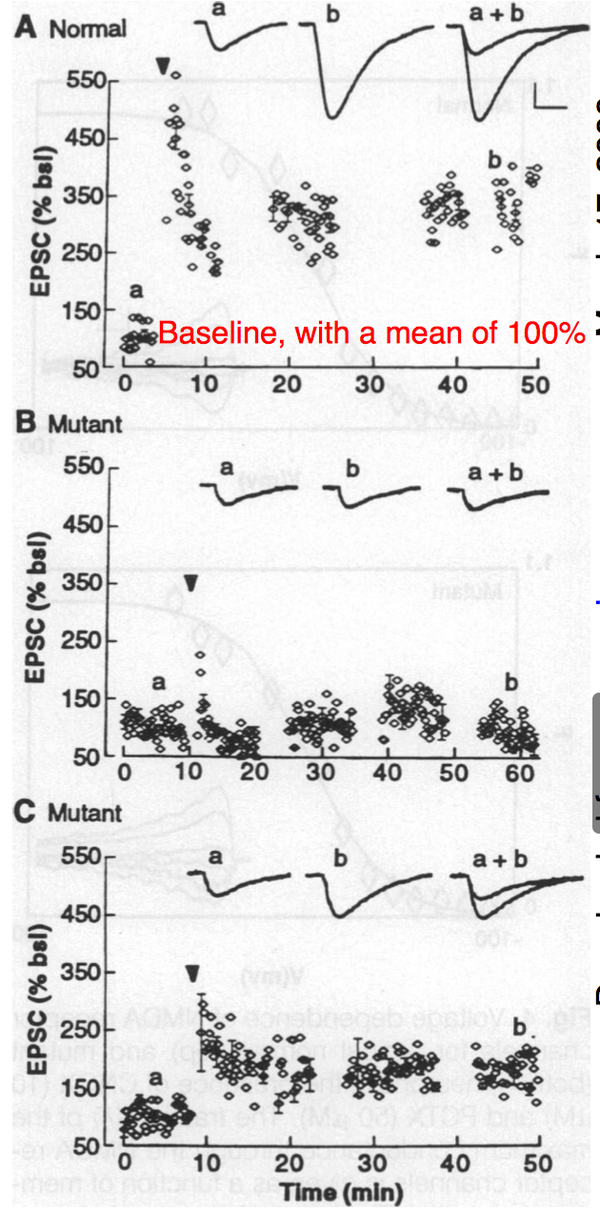

结论:αCaMKII变异小鼠具有明显的LTP缺陷。
二、αCaMKII蛋白上T286位点的自我磷酸化对LTP和学习记忆是否必要?(Giese et al., 1998)
时间已经行进到1998年了。对,就是《相约九八》的那个九八年。小时候听歌,总以为是《相约酒吧》,想不明白为什么1998年就要去酒吧,感觉很不"正经"。
言归正传。
还记得我们一直说的吗,αCaMKII蛋白一旦自我磷酸化之后就不再依赖于Ca++或者CaM的水平。那么自我磷酸化是否是必要的呢?这篇文章要验证的就是这一点。虽然一作换了人,但是通讯作者还是Silva。文章还是发在Science上。
研究模型的建立:
将286位点上的苏氨酸(T286)替换为丙氨酸(T286A),这样一来,该位点就不能转换到不依赖于CaM水平的状态了。也就是说它无法自我磷酸化了。接下来用Cre-LoxP系统进行DNA重组置换,将修改后的序列用质粒载体插入原基因序列中,再用新霉素进行筛选,最后用Cre重组酶剪掉LoxP位点及neo基因,得到目标基因序列。
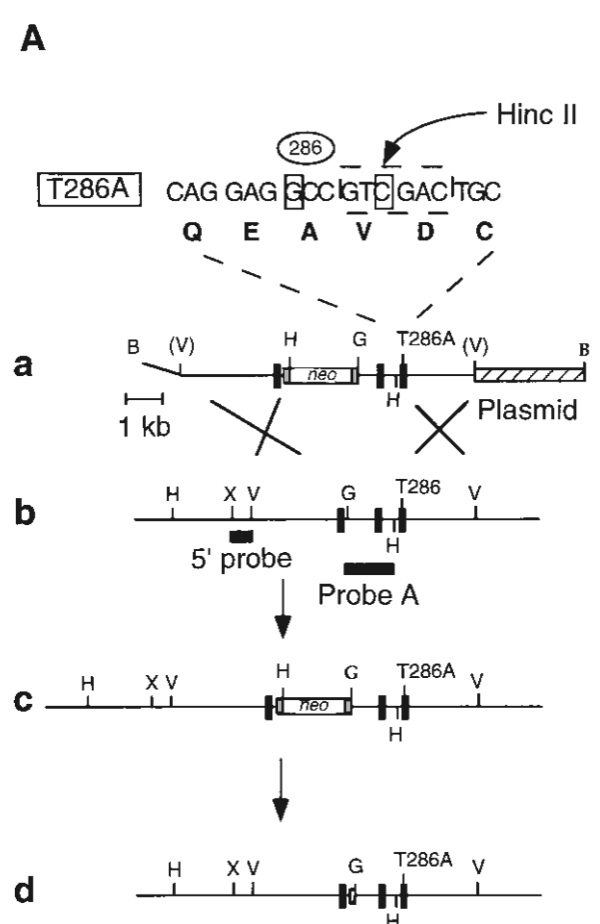
上述模型通过了PCR和Western blot检测,解剖形态学分析也表明,野生型小鼠与变异小鼠的海马区脑组织切片无异。
场电位记录结果,与野生型相比,失去自我磷酸化能力的T286A小鼠没有LTP,但其突触传递能力没有受损。
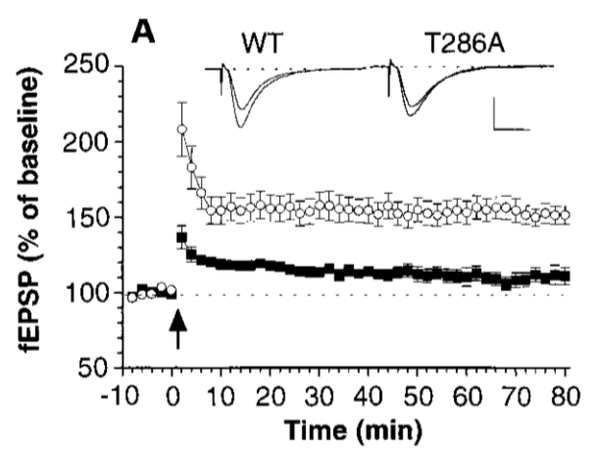
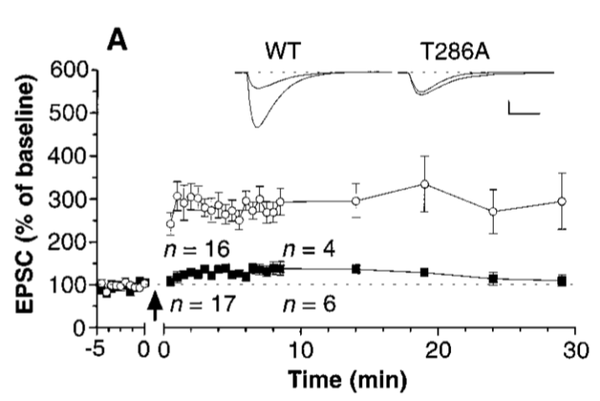
单个神经元的全细胞记录也出现了相似的结果。
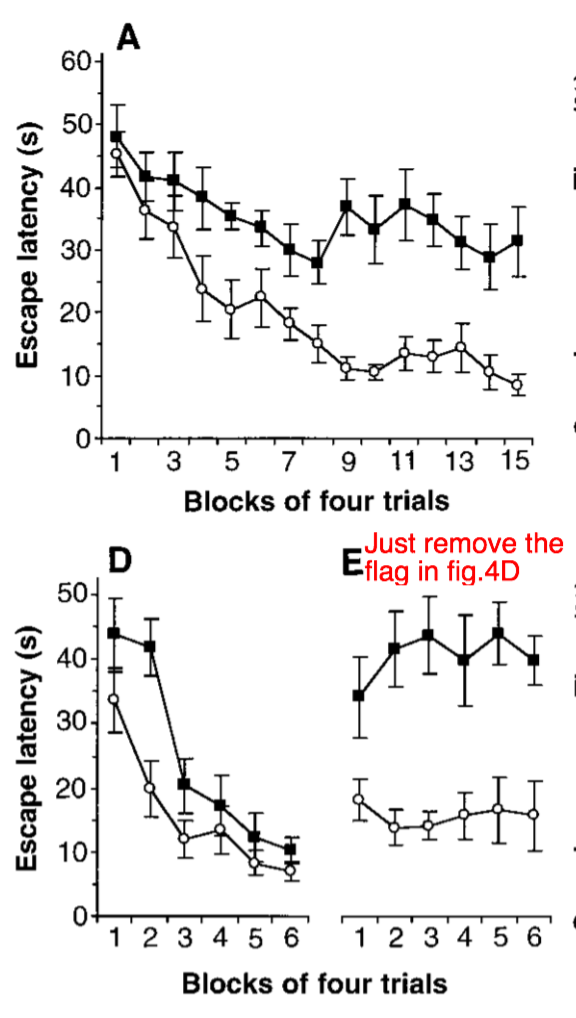
行为学测验用的仍然是莫里斯水迷宫。图中的空心圆圈表示的是野生型的测试结果,而实心方块表示的是变异小鼠的测试结果。可以看到,经过10次左右的测试后,野生小鼠很快就能找到水中的隐藏平台,而变异小鼠需要花费更多的时间(A),并且一旦撤掉可见平台上的旗子,野生小鼠就很难找到平台(D-E)。
结论:αCaMKII蛋白上T286位点的自我磷酸化对LTP和学习记忆来说是必要的。
三、加强αCaMKII的自我磷酸化能够使小鼠变聪明吗?(Mayford et al., 1996)
仍然是Science上的文章。该文发表于1996年。一作又换了人,但是仔细一看,通讯作者是诺奖得主Eric R. Kandel,那本神经科学圣经Principles of Neural Science的编者之一。
在这项研究中,他们采用了与前两篇文章不一样的基因编辑手段,转基因技术。
相比于基因打靶技术,转基因技术不要求进行基因序列定位,只要把载体打进去就好了。这样一来,又省钱省力,但缺点就在于,你不知道转基因究竟转入了原始基因序列中的什么位置。它有可能会干扰正常的基因表达,也可能不会。因此,如果要使用转基因技术进行研究,需要用多个不同的品系,得到一致性结果才能验证你想要验证的结论。
研究者使用了强力霉素(Doxycycline)来调控转基因的表达。如图,他们生产了两种类型小鼠。第一种类型的小鼠能在αCaMKII启动子的调控下表达tTA,而另一个类型的小鼠会在tet-O启动子的调控下表达CaMKII-Asp286。 Asp286是286位点的另一种变异,与上文中无法自我磷酸化的T286A相反,它能使CaMKII上的酶一直处于不依赖于Ca++的状态,也就是一直都在自我磷酸化。
而这两种转基因小鼠交配产生的子代小鼠,部分会同时带有两种基因。这会产生什么样的效果呢?当tTA结合在tet-O启动子上的时候,CaMKII-Asp286就能够表达,但如果给予强力霉素(Doxycycline,DOX),那么DOX就会与tTA结合,并脱离tet-O启动子,从而关闭CaMKII-Asp286的表达。
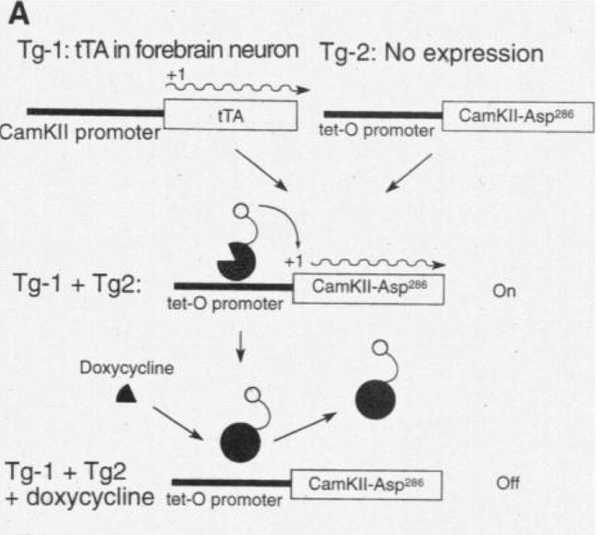
这么做是要干嘛?上一篇文章不是说阻断自我磷酸化小鼠就变笨了不会学习了么。好嘛,那我就让它一直一直磷酸化,是不是就会变聪明啦?这idea真的很棒棒哦。让我们来看看场电位记录和行为学测试的结果。
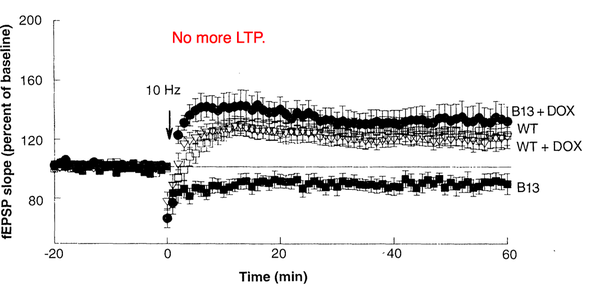
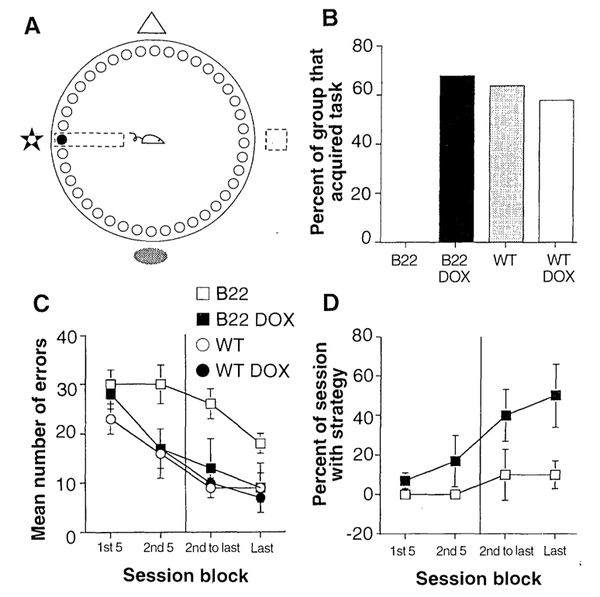
超乎想象的是,一直自我磷酸化并不会产生更强的LTP,反而失去了LTP。并且在空间记忆实验的Barnes水迷宫中,实验老鼠也表现得很差劲(B13和B22表示的是用于转基因的不同品系)。也就是说,过度的自我磷酸化并不能让小鼠变
结论:过度的自我磷酸化并不能增强记忆能力。
四、对记忆力增强的不懈追求 (Tang et al., 1999)
1998~1999年,Nature发了两篇文章,都是关于记忆力增强的实验。1998年的那篇文章(Migaud et al., 1998)表明,通过PSD-95来增强LTP并不能提升记忆力表现。99年的这篇文章则表明,NMDA受体亚型NR2B的国度表达,有利于提高NMDA受体的活性,并促进了突触增强作用。
他们的做法并不复杂,使用的也是转基因技术,让NR2B过表达。
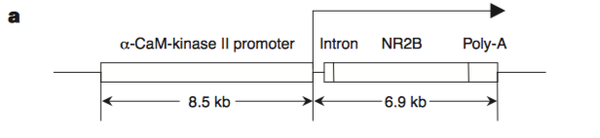

NR2B受体通道的开放时间要长一些。
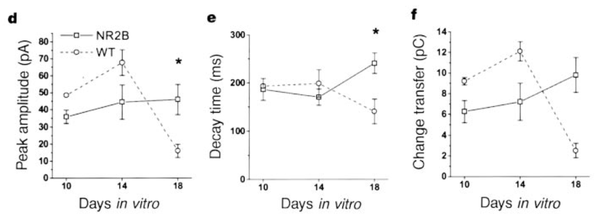
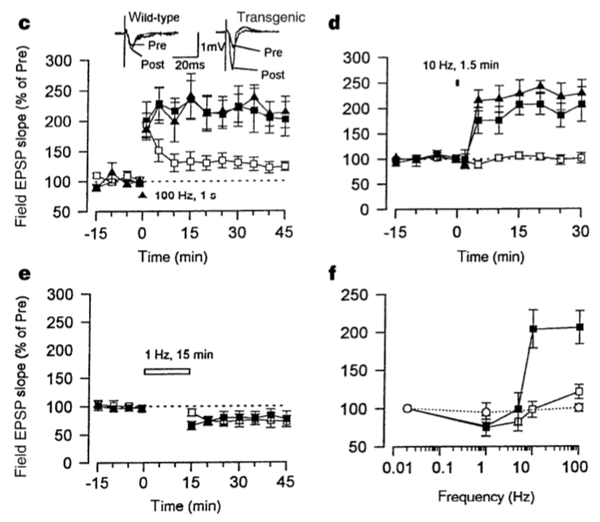
场电位记录表明,NR2B过表达的变异小鼠不仅拥有增强的LTP,而且也保留了LTD。
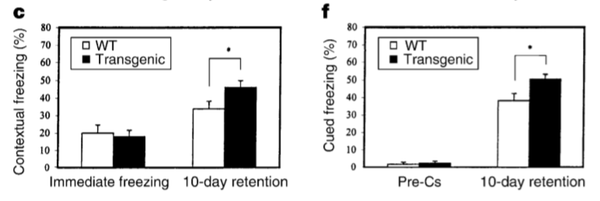
也许正是因此,它们在行为学的测试中才表现得更聪明,恐惧条件发射建立和消退得都更快。
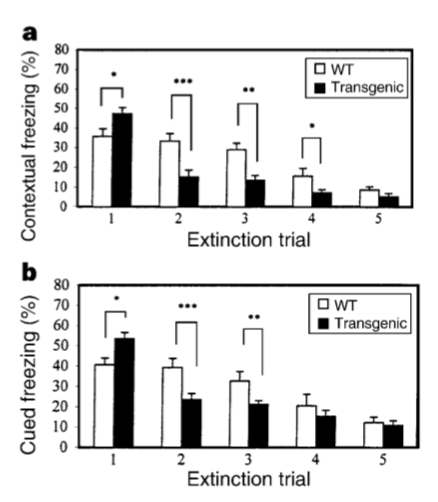
结论:NR2B受体在前脑的过表达能够提高NMDA受体活性,提高突触促进作用,并使个体在学习和级以上的表现更好。
五、结语
时至今日,学习与记忆的研究仍然属于热点问题。我国的脑科学研究重点就在于学习、记忆与教育。为什么学习与记忆这么重要?站在普通年轻人的角度上看,提高记忆力好像无非就是记东西更好一些罢了,貌似也没有什么很大的提升。而且说实话,又能提升多少呢。但是对于某些人群来说,比如智力障碍(intellectual disorder)患者或是老年人,这一点点改变所带来的影响是巨大的,对生活质量的提升也是很明显的。
另外,在接下来的专栏文章中,我们也会提到,记忆与情绪的关联作用。从某种意义上来说,很多情况下,记忆无意识地影响了我们的知觉、情绪、思维、执行能力等各个方面。它好像是你最不可或缺的工具书,最常用的代码模块,最长陪伴于身边的物件,比如一副耳机,一块手表,一支钢笔,而你每天都在无数次地使用它。它与你紧密关联,说它就是你的影子也不为过。
参考文献:
Silva, A. J., Paylor, R., Wehner, J. M., & Tonegawa, S. (2009). Impaired Spatial Learning in L-Calcium-Calmodulin Kinase Mutant Mice, 257, 6.
Silva, A. J., Stevens, C. F., Tonegawa, S., & Wang, Y. (2009). Deficient Hippocampal Long-Term Potentiation in o-Ca cium- Calmodulin Kinase Mutant Mice, 257, 6.
Giese, K. P. (1998). Autophosphorylation at Thr286 of the Calcium-Calmodulin Kinase II in LTP and Learning. Science, 279(5352), 870–873. https://doi.org/10.1126/science.279.5352.870
Migaud, M., Charlesworth, P., Dempster, M., Webster, L. C., Watabe, A. M., Makhinson, M., … Grant, S. G. N. (1998). Enhanced long-term potentiation and impaired learning in mice with mutant postsynaptic density-95 protein. Nature, 396(6710), 433–439. https://doi.org/10.1038/24790
Tang, Y.-P., Shimizu, E., Dube, G. R., Rampon, C., Kerchner, G. A., Zhuo, M., … Tsien, J. Z. (1999). Genetic enhancement of learning and memory in mice. Nature, 401(6748), 63–69. https://doi.org/10.1038/43432
来源:知乎 www.zhihu.com
作者:知乎用户(登录查看详情)
【知乎日报】千万用户的选择,做朋友圈里的新鲜事分享大牛。 点击下载
没有评论:
发表评论