登上陆地,还是继续留在水里——对石炭纪的四足动物来说,这是一个问题。
它们继承了泥盆纪祖先的演化成果:两套并行的生存装备。在水下用鳃(还有皮肤)呼吸,用尾巴划水;上岸后用肺(还有皮肤)呼吸,用四肢慢慢爬行。石炭纪茂密的雨林和沼泽成为这支两栖部队的主战场。
但是水下和陆地毕竟是两种完全不同的生境。水和空气的物理化学性质:密度,浮力,阻力和推进力,还有折射率和氧含量天差地远。对浸没于其中的任何动物,都不可能有一套完美的身体构型,让它们在水下矫若游龙,还能在陆地上敏如脱兔。而且同时供养两套装备也是个不小的负担,加上食物和天敌的区别。于是不可避免地,早期四足动物们开始向着强化水栖功能和强化陆栖分头演化。
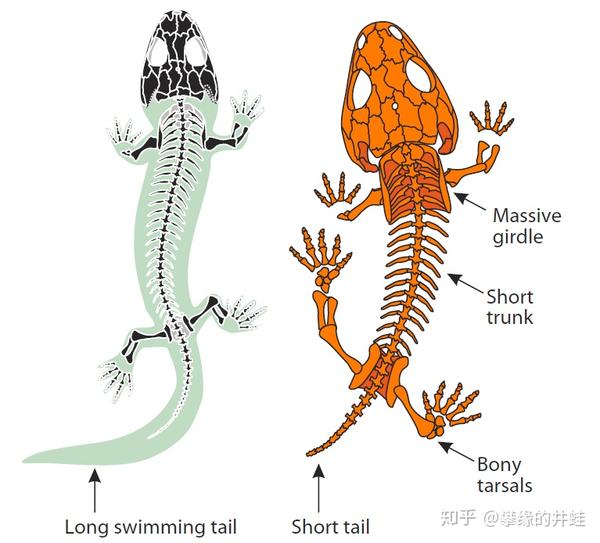
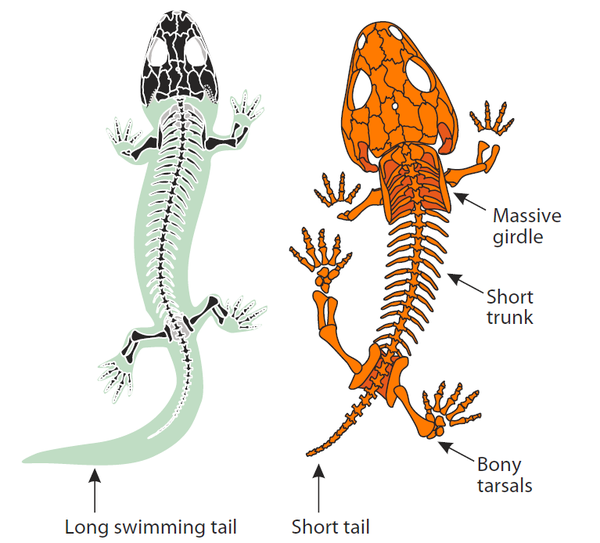
两种早期四足动物身体结构。左:Micromelerpeton,晚石炭世,水栖;右:Micropholis,早三叠世,陆栖。水栖的种类身体更符合流体力学,有宽扁的大尾巴用来推进。陆栖的种类身体粗壮;颅骨,脊柱,以及四肢和肢带的骨骼更加强劲。尾巴变得短小,减少在地面拖曳带来的阻力。图片来源自[1]。

现代蓝舌石龙子(Tiliqua scincoides)爬行时前肢(上图)和后肢(下图)骨骼的运动方式。随着肢体的屈伸,脊柱也左右扭曲成S形。石炭纪的陆栖四足动物,比如彼得足螈和引螈,很可能就是用这种方式运动。图片来源自[2]。
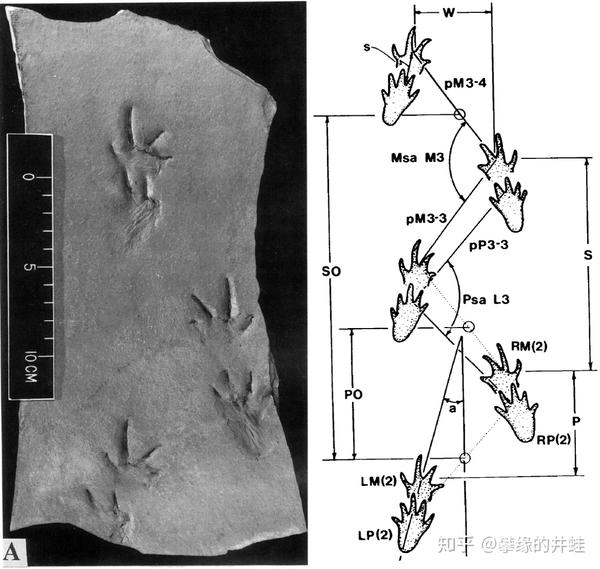
晚石炭世陆栖四足动物的足迹化石。当它扭动身体时,弯曲的一侧前脚和后脚几乎碰到一起,留下了好像叠在一起的两个脚印。图片来源自[3]。
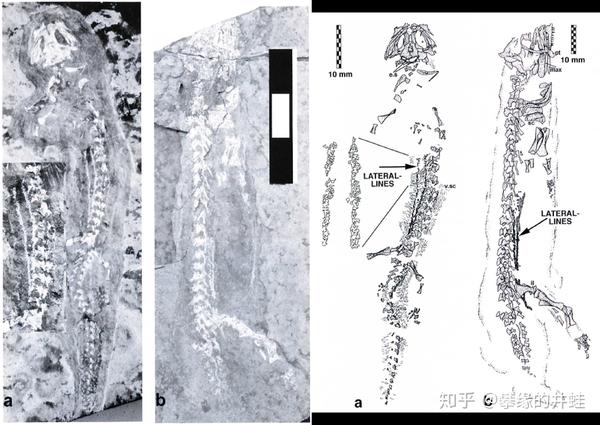
两种晚石炭世到早二叠世离片椎类:(a)幻螈Apateon pedestris;(b)Melanerpeton humbergenses。在它们的化石上找到了侧线(Lateral line)的痕迹。侧线是感知水中细微震动的器官【参见第一百五十五回 起跑线】,说明这两种动物是水生的。陆生四足动物侧线消失,演化出中耳(Middle ear)来接收和放大空气的震动。图片来源自[4]。
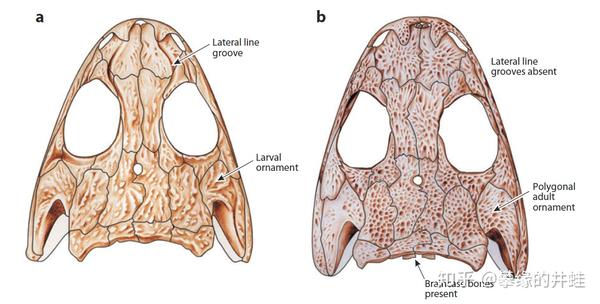
在骨骼化石上也可以找到一些蛛丝马迹。(a)是Micromelerpeton幼体的头骨,上面可以观察到侧线槽(Lateral line groove),是侧线下方神经生长的位置;(b)是Micromelerpeton成体的头骨,侧线槽消失,骨骼结构也发生了变化。推测Micromelerpeton会随着生长,生活习性由水栖逐渐变为偏陆栖。图片来源自[1]。
水栖还是陆栖的抉择在非羊膜类四足动物的生活史中集中体现。现代两栖动物有一个独特的幼体阶段:蝌蚪。那么石炭纪的大家伙呢?难道也是从蝌蚪变来的吗?幸运的是,有一些早期四足动物留下了非常翔实的化石资料,包括不同发育阶段的幼体,让我们得以一窥它们的生命周期。
下面是一些石炭纪离片椎类Sclerocephalus haeuseri【参见第一百九十一回Fish
to Beast】的化石,详细地展示了这种动物的发育过程(图片全部来源自[5]):

刚刚孵化不久的幼体(标尺长度1厘米)。大脑袋,四肢纤弱,身体的骨骼还没有发育完全,运动能力很差,正处在最脆弱的阶段。
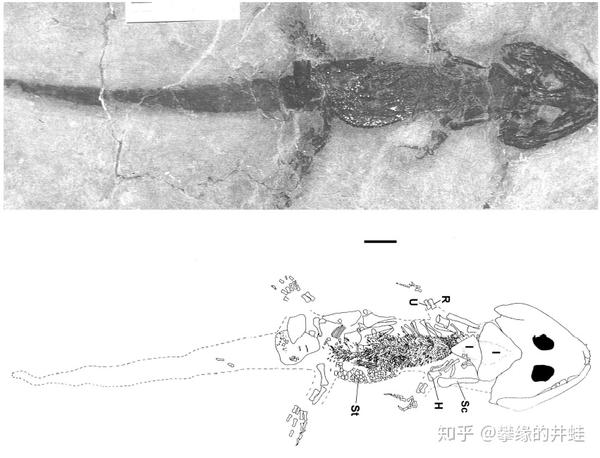
长大一些的幼体(标尺长度1厘米),脊椎,肋骨和肢骨发育成型,身体和尾巴的比例增大,捕食和逃生的能力增强。

再大一些的亚成体(标尺长度5厘米)。全身骨骼已经非常坚实,配上发达的肌肉,已经跻身中等掠食者的行列。

成体体长超过1米(标尺长度20厘米),头骨扁长,口阔齿利,凶猛贪婪,是水滨一霸。化石下方前后肢中间的是腹部鳞片的印痕。和滑溜溜的现代两栖类不同,S. haeuseri的身体上(至少肚皮上)覆盖着细密的鳞片。
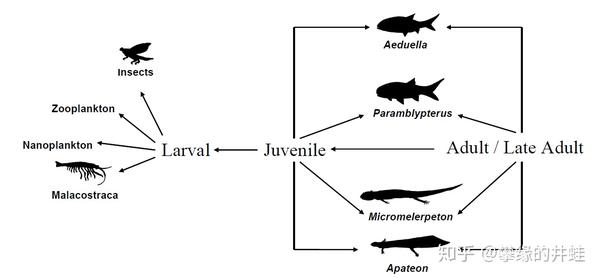
不同发育阶段的S. haeuseri位于食物链的不同等级。幼体只能吃一点浮游生物,节肢动物;半成体和成体的猎物包括水域里的各种鱼类和四足动物,还有小一点的同类。
S. haeuseri的发育过程除了体型增大,骨骼发育,身体比例的调整之外,还有一个非常重要的变化。
S. haeuseri的早期幼体,头部两侧有一些延伸出来的羽状结构:
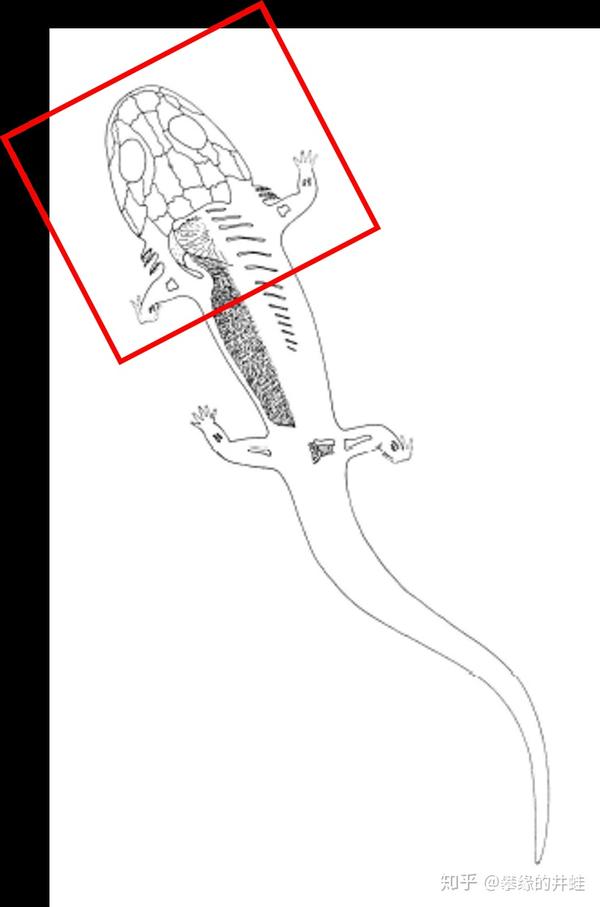
如果放大来看,是这样的:
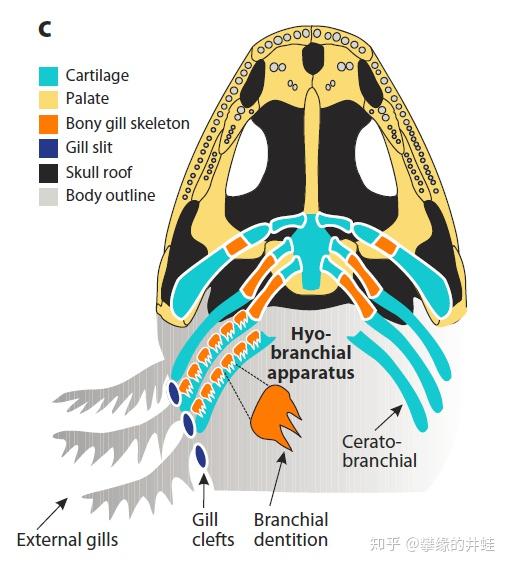
石炭纪非羊膜类四足动物幼体具有外鳃(External gills)作为呼吸器官。还有鳃裂和软骨质的鳃笼,上面生有鳃齿(Gill teeth,Branchial Dentition),用来钉牢和撕裂食物。这都是和水生生活相适应的特征。图片来源自[1]。
随着身体发育,S. haeuseri的肺逐渐长成并承担呼吸功能,外鳃萎缩消失,鳃笼和鳃齿也被更加坚固的骨骼替代。整个发育过程就像这样:
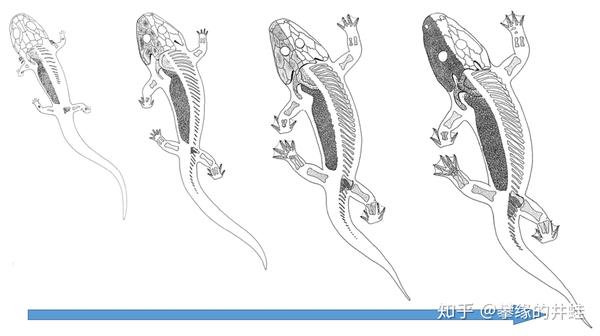
大多数石炭纪的非羊膜类四足动物都是这样长大的,S. haeuseri的成体营水陆两栖生活,保留了流线形的身体和大尾巴。而像引螈这样成体高度陆栖的种类,幼体和成体的体形差异会更加明显。
另一类生活在石炭纪到二叠纪之交的离片椎类四足动物:鳃龙类(Branchiosauridae)也留下了丰富的化石资料。顾名思义,许多鳃龙类终生水栖,成体也会保留外鳃。这种现象被称为"幼态持续(Neoteny)"。有些鳃龙很可能是其他四足动物的水生幼体,但幻螈属(Apateon)和鳃龙属(Branchiosaurus)的发育过程已经研究得非常清楚了。

一些早二叠世鳃龙科化石。(a)幻螈Apateon dracyiensis;(b)鳃龙Branchiosaurus salamandroides; (c)一群幻螈Apateon dracyiensis。鳃龙类体型小巧(标尺长度1厘米),经常沦为大个子近亲(比如S. haeuseri)的口中餐。图片来源自[6]。
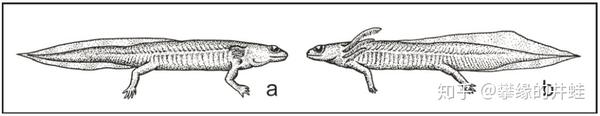
鳃龙类复原图。同一种类的鳃龙在不同的生活环境中形态也会出现明显的差异。生活在溶解氧丰富的溪流中的个体(a)外鳃短,尾巴修长;而生活在氧含量较低的静水(湖沼和池塘)中的个体(b)外鳃发达,尾鳍宽大。图片来源自[6]。
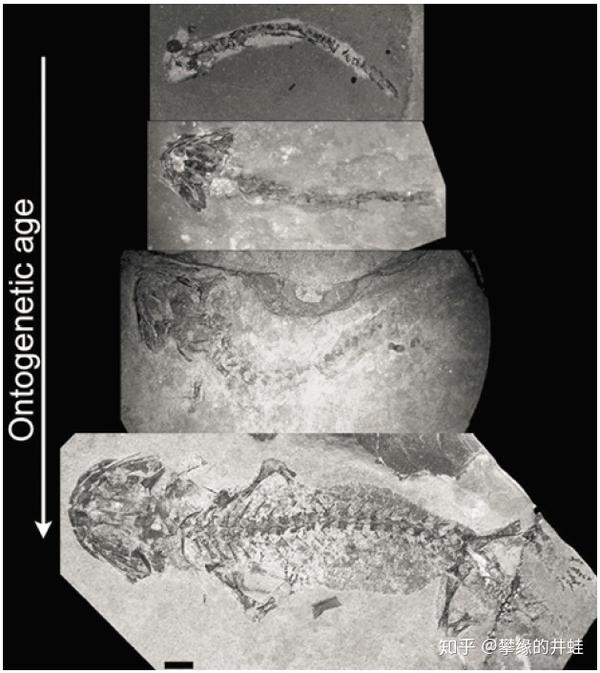
不同发育阶段的幻螈化石。最上方的是刚刚孵化的幼体,没有硬骨也没有四肢。之后颅骨首先骨化,然后是脊柱,之后再长出四肢。图片来源自[7],标尺长度2毫米。
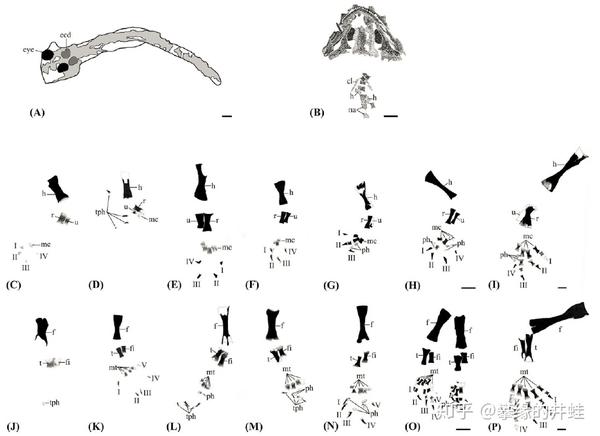
幻螈四肢(前肢C-I;后肢J-P)的发育过程,A是没有四肢的早期幼体,B刚刚萌发肢芽。图片来源自[7],标尺长度1毫米。
对比一下S. haeuseri和幻螈:S. haeuseri的早期幼体具有成型(虽然很弱小)的四肢,除了外鳃,形态和成体差别不大;而幻螈幼体有相当长的阶段没有四肢,完全不像成体——这就是我们非常熟悉的蝌蚪——和昆虫几乎同时,四足动物也演化出了变态发育(Metamorphosis)。
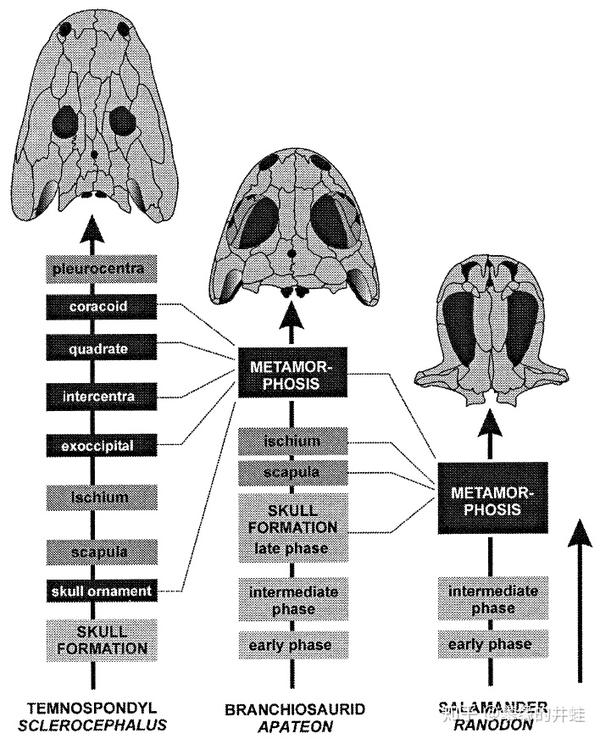
S. haeuseri(左)和幻螈(中)的发育过程对比。S. haeuseri可以称为"无变态",幼体渐次发育为成体。而幻螈在蝌蚪阶段的末期,一系列骨骼(鸟喙骨Coracoid,方骨Quadrate,间椎Intercentra,枕外骨Exoccipital)快速生长,颅骨重排,四肢成型,在短时间内变态发育为亚成体。变态发育可以使幼体和成体占据完全不同的生态位,减少种内竞争,提高环境适应性。现代两栖类(右,Ranodon北鲵属)进一步发展了变态过程。图片来源自[8]。
现在可以总结一下早期非羊膜类四足动物的发育过程了:它们的卵在水中孵化,早期的幼体生活在水里,用鳃呼吸,用尾巴游泳,有四肢或者没有四肢(蝌蚪)。有四肢的幼体慢慢长大,它们中的一些离开水体,到陆地上生活,只在繁殖时才回到水中;另一些则终生水栖;当然,也有游走于水陆之间的种类。没有四肢的蝌蚪长大后,短时间内变态发育成成体,登上陆地或者呆在水里。
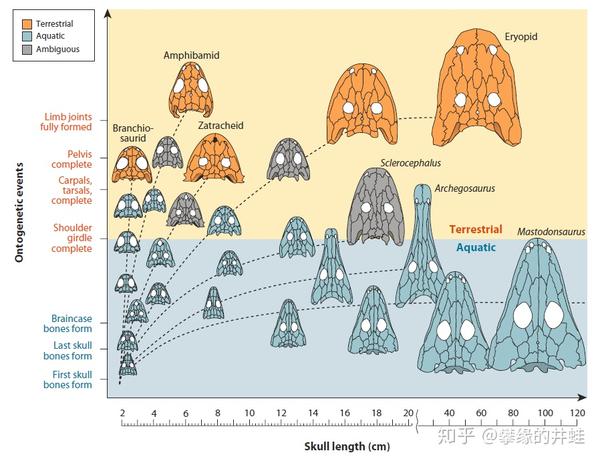
一些离片椎类的发育过程。蓝色的是水栖的幼体和成体,紫色的是两栖生活的半成体和成体,橙色的是陆栖的成体。图片来源自[1]。

一只幻螈Apateon gracilis幼体和它肚子里(红框内)的最后一餐:一只更小的同类。虽然体型纤小,鳃龙类也是凶残的猎手。图片来源自[9]。
地球名片
生物分类:动物界-脊索动物门-脊椎动物亚门-四足形类-离片椎类-鳃龙科
存在时间:石炭纪晚期 至 二叠纪末期
现存种类:无
化石种类:不明
生活环境:淡水,陆地
代表特征:体小,骨骼纤弱,许多种类幼态持续,成体也保留外鳃
包含种类:鳃龙,幻螈
参考文献:
[1] Rainer R. Schoch, Evolution of Life Cycles in Early Amphibians. Annu. Rev. Earth Planet. Sci. 2009. 37: 135–62, doi: 10.1146/annurev.earth.031208.100113
[2] John A. Nyakatura, Emanuel Andrada, Stefan Curth, et al., Bridging'Romer's Gap': Limb Mechanics of an Extant Belly-Dragging Lizard Inform Debate on Tetrapod Locomotion During the Early Carboniferous. Evol. Biol., DOI: 10.1007/s11692-013-9266-z
[3] Frederick A. Sundberg, J Bret Bennington, Michael C. Wizevich, et al., Upper Carboniferous (Namurian) amphibian trackways from the Bluefield Formation, West Virginia, USA. Ichnos, v. 1, p. 111-124, 1990
[4] Ralf Werneburg, Schleusingen, First record of postcranial lateral lines in fossil amphibians (Branchiosauridae; Rotliegend Upper Carboniferous – Lower Permian). N. Jb. Geol. Paläont. Abh., 232(2/3), 365-373, 2004
[5] Ulla Lohmann, Sven Sachs, Observations on the Postcranial Morphology, Ontogeny and Palaeobiology of Sclerocephalus haeuseri (Amphibia: Actinodontidae) from the Lower Permian of Southwest Germany. Memoirs of the Queensland Museum 46(2): 771-781
[6] Schneider, J.W. & Werneburg, R. (2012): Biostratigraphie des Rotliegend mit Insekten und Amphibien. – In: Deutsche Stratigraphische Kommission (Hrsg.; Koordination und Redaktion: H. Lützner & G. Kowalczyk für die Subkommission Perm-Trias): Stratigraphie von Deutschland X. Rotliegend. Teil I: Innervariscische Becken. –
Schriftenreihe der Deutschen Gesellschaft für Geowissenschaften, Heft 61:
110-142; Hannover.
[7] Nadia B. Fröbisch, Ontogeny and phylogeny of small dissorophoid amphibians. A thesis submitted to McGill University in partial fulfillment of the requirements of the degree of Doctor of Philosophy
[8] RAINER R. SCHOCH, NADIA B. FRÖBISCH, METAMORPHOSIS AND NEOTENY: ALTERNATIVE PATHWAYS IN AN EXTINCT AMPHIBIAN CLADE. Evolution, 60(7), 2006, pp. 1467–1475
[9] Florian Witzmann, Cannibalism in a small growth stage of the Early Permian branchiosaurid Apateon gracilis (Credner, 1881) from Saxony. Fossil Record 12 (1) 2009, 7–11 / DOI 10.1002/mmng.200800006
感兴趣的话,请关注我的微信公众号:攀缘的井蛙。每天琢磨点新东西。
来源:知乎 www.zhihu.com
作者:攀缘的井蛙
【知乎日报】千万用户的选择,做朋友圈里的新鲜事分享大牛。 点击下载
没有评论:
发表评论