
自诞生的一刻起,地球上所有的生命就被投入了一场永无休止,而且不断加速的生存竞赛,争夺的焦点就是有限的资源,配偶和生存空间。异养生活的动物们主要拼军备,用力量,速度,智慧,杀伤力和繁殖力压倒对手;而作为自养生物,植物的竞争主要是靠拼基建,一切能最大限度地保障光合作用进行,提高反应效率的性状都会被保留和强化,最明显的就是营养器官(根茎叶),尤其是叶片的出现和分化。
最早登上陆地的维管植物先驱(参见第一百十二回 芽)没有叶子,用裸露的地上茎完成光合作用。
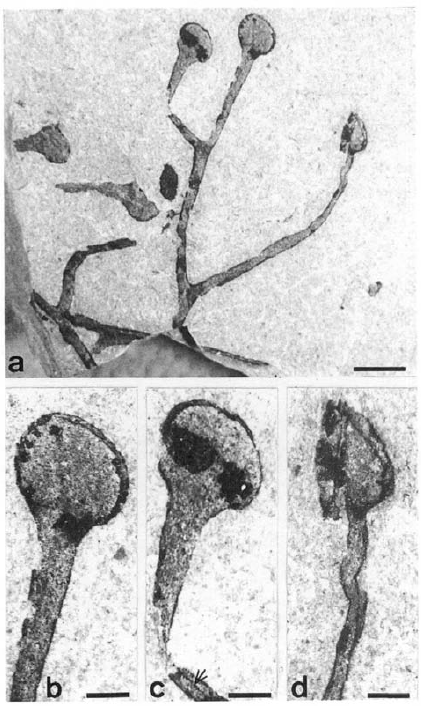
志留纪顶囊蕨Cooksonia cf caledonica全株(a,标尺长度2.1毫米)和孢子囊(b-d,标尺长度0.5毫米)化石。地上茎只是一丛秃秃的光杆,杆头顶着膨大的孢子囊。图片来源自[1]。
但是茎的结构并不适合这项工作。当原本的不毛之地被越来越多的植被覆盖,对阳光的争夺战迫使植物改良身体结构,茎专注于支撑功能,变得越来越粗壮强韧,也越来越粗糙灰暗;气孔和叶绿体转移集中到茎顶端的小枝上,为了提高受光面积,这些小枝变得扁平宽大,内部贯穿维管系统,源源不断地运输原料和成品,成为专门的光合工场。于是,最早的叶片诞生了。
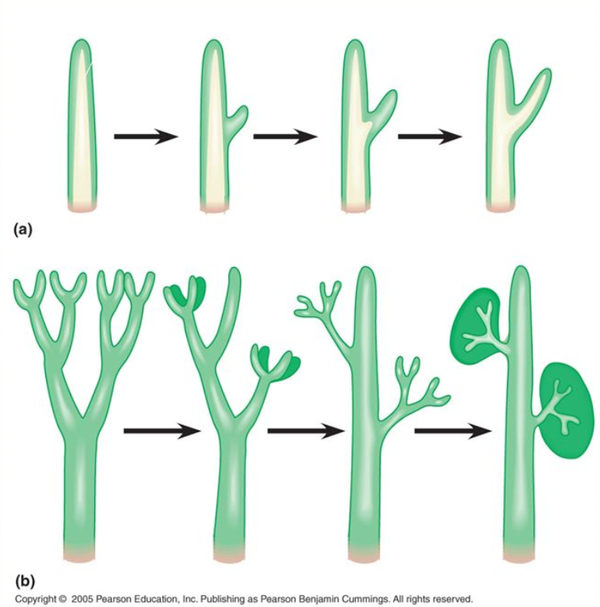
叶片产生的两种假说。(a)茎表突起维管化;(b)侧枝扁化合并。图片来源见水印。
目前认为石松植物的叶片是通过茎表突起维管化(突起说)产生的,这种叶片结构简单,内部只有一条不分枝的单一叶脉,称为小型叶(Microphylls)。几乎在同一时间,还有一批维管植物(真叶植物,Euphyllophyte)通过侧枝扁化合并(顶枝说)独立演化出另一类叶片,这类叶片具有复杂的分枝叶脉,称为大型叶(Megaphylls)。【感谢 @兔小灰 在植物叶的进化历程是怎么样的?的回答】
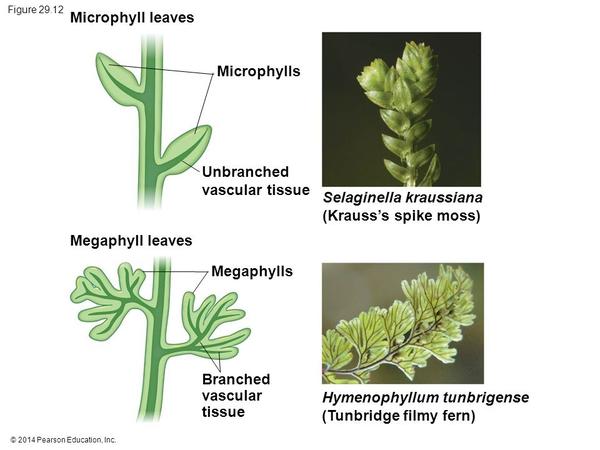
小型叶(上,实例是小翠云卷柏Selaginella kraussiana)和大型叶(下,实例是汤氏膜叶蕨Hymenophyllum tunbrigense)结构对比。图片来源见水印。

现代石松的小型叶放大图,在茎上细密排列。古生代的木本石松也长着这样的小型叶,叶片脱落后,就会在茎的表面留下鳞片状的细密叶座(参见第一百八十四回 林网)。图片来源自网络。
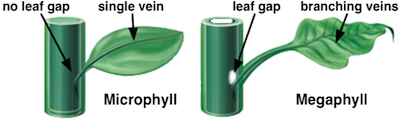
小型叶(左)和大型叶(右)的区别并不在大小,而在于结构。和大型叶相比,小型叶还缺少一个叫做"叶隙(leaf gap)"的结构。图片来源自网络。
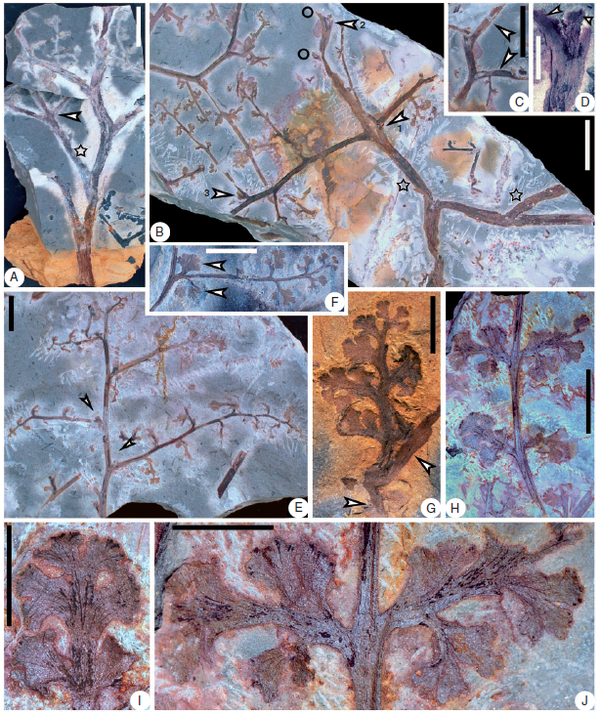
早泥盆世守刚蕨Shougangia bella枝条和片状叶化石和结构图。代表着真叶植物和大型叶的演化早期形态。图片来源自[2]。
真叶植物包括节蕨类,真蕨类,以及种子植物。需要指出的是,不同类群的大型叶很可能是各自独立起源的。在石炭纪,真叶植物的多样性已经非常高,留下了非常丰富的化石资料。
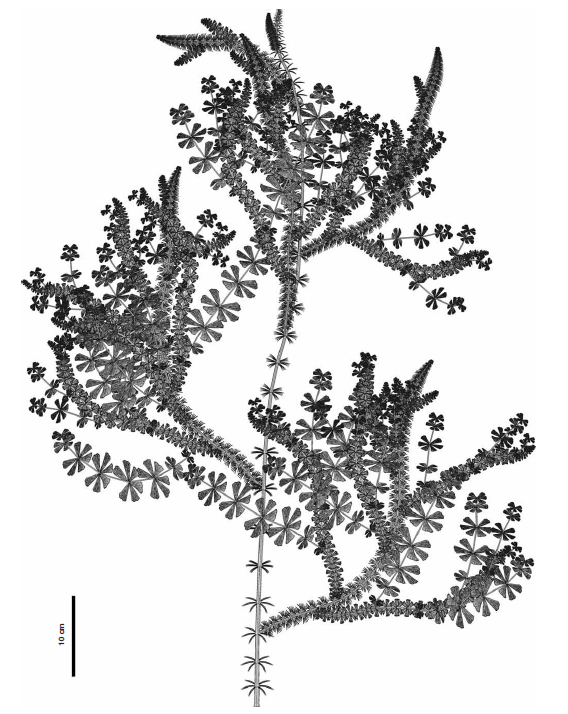
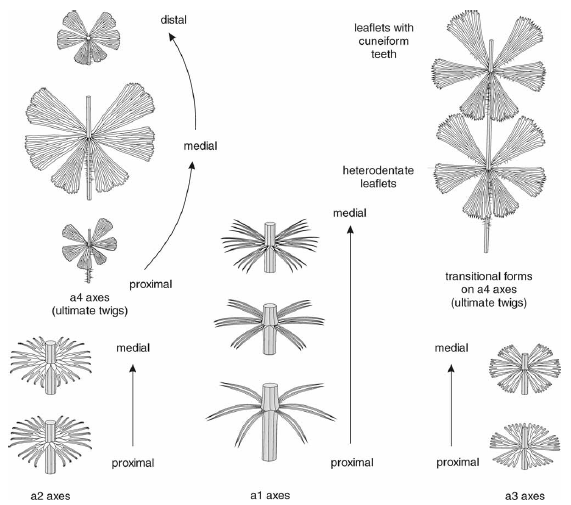
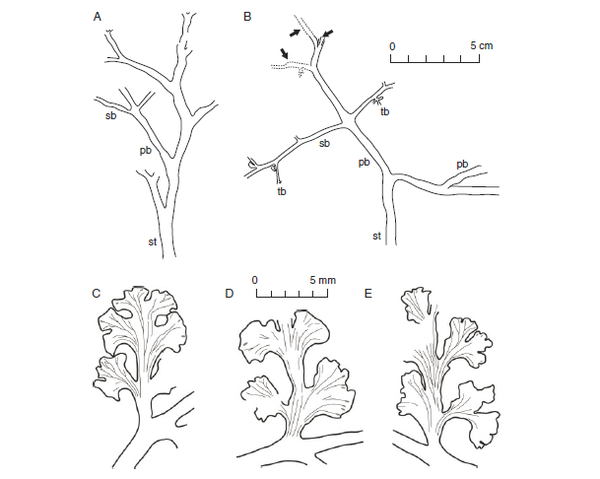
石炭纪轮叶Sphenophyllum costae的复原图(参见上回),不同部位的叶片形态差别很大。基部主干基本接触不到阳光,叶片趋于退化,变成细小的针状;植株上方的分枝是主要的采光部位,上面的叶片成宽大的扇形,是光合生产的主力。图片来源自[3]。
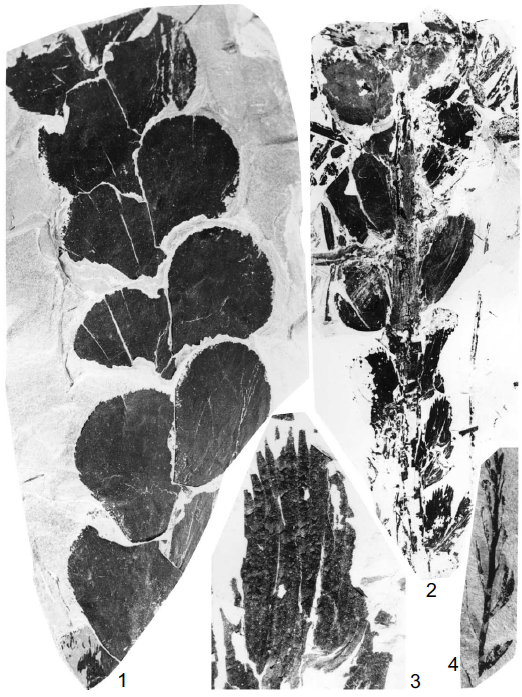
瓢叶目(Noeggerathiales)生活在古生代晚期,这类植物的系统分类一直不明。它们的大型叶基部抱合在茎轴上,同一株植物上也生有多种形态的叶片。上图是石炭纪瓢叶类Noeggerathia foliosa化石。 图片来源自[4]。
晚石炭世到早二叠世的真叶植物化石中,数量最多,形态最多样的,是真蕨类(Polypodiopsida)和种子蕨(Pteridospermatophyta)。这两类植物的叶片十分相似,都具有特征明显的复杂羽裂,经常被混称为羊齿植物,属名中多带有词根"pteris",比如畸羊齿 Mariopteris,扇羊齿 Rhacopteris,楔羊齿 Sphenopteris,栉羊齿 Pecopteris,脉羊齿 Neuropteris,齿羊齿 Odontopteris等等。下面是一些羊齿植物的叶片化石:

两种石炭纪畸羊齿Mariopteris muricata(5)和Mariopteris dernoncourtii(6)叶片化石。图片来源自[5]。

石炭纪栉羊齿Pecopteris nyranensis(3)叶片化石。图片来源自[5]。

石炭纪楔羊齿Sphenopteris germanica叶片化石,图片来源自[6],标尺长度1厘米。

石炭纪齿羊齿Odontopteris schlothemii化石。图片来源自[6],标尺长度1厘米。
石炭纪脉羊齿Neuropteris ovata化石。图片来源自[6],标尺长度1厘米。
这样的叶子在今天也不鲜见。种子蕨虽然灭绝已久,但10000多种现代真蕨类依然保持着这种古老的叶型。由于造型古拙典雅,许多蕨类都被培育成观赏植物,不用特地到山野林地中去找寻,也能在城市花房里找到它们的身影。
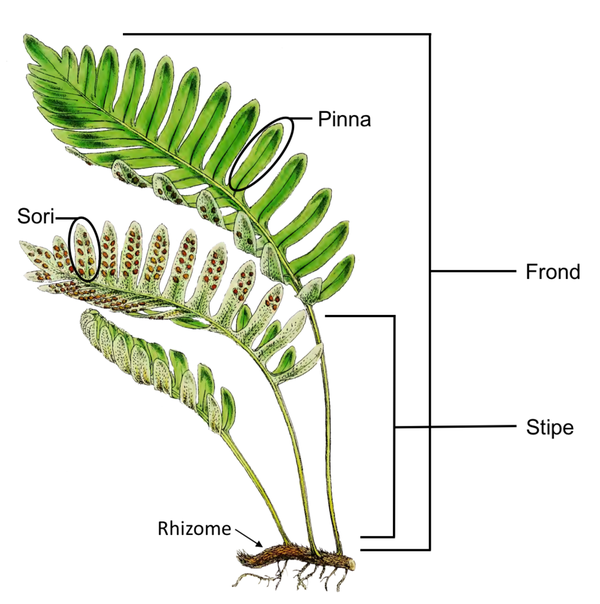
现代真蕨类植物结构,蕨叶(Frond)由地下茎(Rhizome)抽出,基部是细长的叶柄(Stipe),叶片大多整齐地分裂成许多细长的羽片(Pinna)。孢子囊(Sori)就生长在生殖叶的背面。图片来源自网络。绝大多数现代真蕨类都是低矮的草本或藤蔓,但在石炭纪,高大的树蕨随处可见。种子蕨的叶片和真蕨类非常相似,但叶片上生长的不是孢子囊,而是种子。
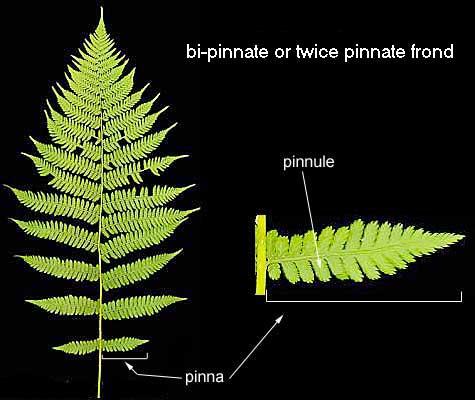
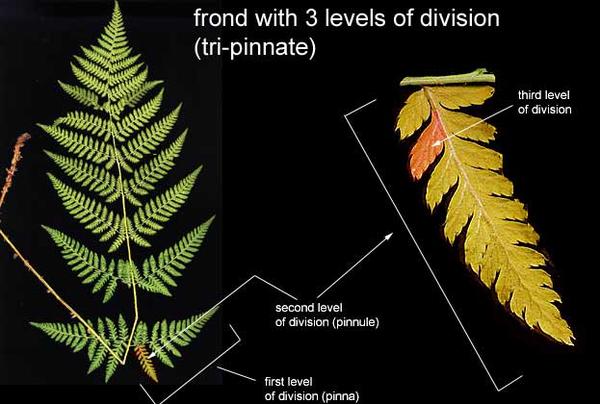
许多蕨类的羽片(Pinna)还会再进一步分裂成更小的Pinnule(小羽片),甚至Pinnule上还有锯齿状的分裂。组成一种具有奇妙美感的分形图像。图片来源自网络。
这样复杂的形状至少有一个好处,那就是适合拼凑,减少了同一株植物叶片之间的覆盖,增大了有效采光面积。通过对叶片大小和生长方向的调节,甚至可以制造出类似"镶嵌"的效果,进一步减少生长浪费,提高光合效率。

现代桫椤目(Cyatheales)树蕨的叶片排列,规整得像是人工修整过一样,相互间错开一定角度生长,插空避让,最大限度地接收阳光。图片来源见水印。
新生的蕨叶蜷曲成类似发条的螺旋形,这种极具特色的嫩芽有一个非常形象的英文名:fiddlehead(提琴头)。

一丛扎根在石缝里的虎尾铁角蕨Asplenium incisum,十几根Fiddlehead正在茁长成长。图片来源自网络。

Fiddlehead近照。每一枚羽片都被精心地盘绕在里面,随着嫩芽的生长从下到上依次舒展开来。蕨类植物没有鲜艳芬芳的花朵,但自有一种清丽秀雅的美。图片来源自网络。

一些蕨类的Fiddlehead被当做鲜嫩的蔬菜,精心烹调后端上餐桌。国内称为蕨菜,井蛙在宣城的小饭店吃过一次,口感清爽滑润。图片来源自网络。

这是一块来自马荣溪的石炭纪动物粪便化石(Coprolites),可以明显辨认出含有羊齿植物的叶片。根据大小推测,它很可能出自一条巨型节胸蜈蚣的肠道。享用蕨叶美餐这事也是古已有之了。图片来源自[7]。
在古生代晚期,羊齿植物也演化出许多高大的木本种类。种子蕨留在后面的章节中介绍,最著名的木本真蕨类当数辉木属(Psaronius),这是一类株高超过10米的高大乔木,和各种石松类,芦木,种子蕨和原始的松柏类混生在一起,是重要的成煤植物。

根据不同部位的化石碎片还原的辉木整株外观。图片来源自网络。

在古生代到中生代,高大的树蕨曾经遍布世界各地,但今天只有桫椤目的少数种类还保持着高大木本的形态。想到引螈,肺蝎和节胸蜈蚣都曾经在这样的蕨伞下游荡歇息,这种感觉真的非常奇妙。图片来源自网络。
更多的真蕨类以低矮的草本或灌木面目出现。它们生长在高大树木的遮蔽之下。今天的蕨类继承了祖先的习性,由于植株矮小,耐阴,常被当做室内盆景。但大多数蕨类对湿度要求很高,不容易养好。

一些石炭纪真蕨类组成的沼泽森林。攀附在辉木树干上的是藤本蕨类Ankyropteris brongnartii ,树下生长的是群囊蕨Botryopteris forensis ,图片左侧是丛生的Zygopteris herryvillensis。图片来源自[8]。
地球名片
生物分类:植物界-真蕨类
存在时间:泥盆纪晚期 至 现代
现存种类:10000种以上
化石种类:不明
生活环境:陆地
代表特征:大型叶,多为羽状或掌状,孢子繁殖
代表种类:桫椤,石韦,贯众,芒萁,骨碎补,井栏边
参考文献:
[1] E. MOREL, D. EDWARDS, M. INIGEZ RODRIGUEZ, The first record of Cooksonia from South America in Silurian rocks of Bolivia. Ceol. Mag. 132 (4). 1995, pp. 449-452
[2] De-Ming Wang, Hong-He Xu, Jin-Zhuang Xue, et al., Leaf evolution in early-diverging ferns: insights from a new fern-like plant from the Late Devonian of China. Annals of Botany 115: 1133–1148, 2015, doi:10.1093/aob/mcv049
[3] Arden R. Bashforth, Erwin L. Zodrow, Partial reconstruction and palaeoecology of phenophyllum costae (Middle Pennsylvanian, Nova Scotia, Canada). Bulletin of Geosciences, Vol. 82, 4, 2007
[4] Zbyněk Šimůfinek, Jiří Bek, Noeggerathiaceae from the Carboniferous basins of the Bohemian Massif. Review of Palaeobotany and Palynology, 125 (2003), 249-284
[5] Stanislav Opluštil, Zbyněk Šimůnek, JosefPšenička, et al., A 25 million year macrofloral record (Carboniferous–Permian) in the Czech part of the Intra-Sudetic Basin; biostratigraphy, plant diversity and vegetation patterns. Review of Palaeobotany and Palynology 244(2017), 241–307
[6] WILLIAM A. DIMICHELE, DAN S. CHANEY, SPENCER G. LUCAS, et al., MIDDLE AND LATE PENNSYLVANIAN FOSSIL FLORAS FROM SOCORRO COUNTY, NEW MEXICO, U.S.A. Lucas, S.G., DiMichele, W.A. and Krainer, K., eds., 2017, Carboniferous-Permian transition in Socorro County, New Mexico. New Mexico Museum of Natural History and Science Bulletin 77
[7] https://paleobiology.si.edu/mazoncreek/index.html
[8] William A. DiMichele, Tom L. Phillips, The ecology of Paleozoic ferns. Review
of Palaeobotany and Palynology 119 (2002) 143-159
感兴趣的话,请关注我的微信公众号:攀缘的井蛙。每天琢磨点新东西。
来源:知乎 www.zhihu.com
作者:攀缘的井蛙
【知乎日报】千万用户的选择,做朋友圈里的新鲜事分享大牛。 点击下载
没有评论:
发表评论