
1.植被类型
在上一篇气候篇中,提到了Wolfe J A(1987)等人对北美地区的植被化石所做的系统研究。研究显示,热带气候可能扩展到了北纬45-50度。因为降水稀少的缘故,植被类型为稀疏的热带常绿阔叶林。在北纬50-65度的范围内,降水增多,植被类型为温带常绿阔叶林。直到北纬65度,常绿植物才被落叶植物所取代。
不过,Golovneva L B(2000)则指出,前者的模型可能高估了降水量,因为植物化石多发现于水源充沛的沉积环境。根据Golovneva L B(2000)的推算,在中纬度地区,离水源较远的植被仍然是稀疏的。
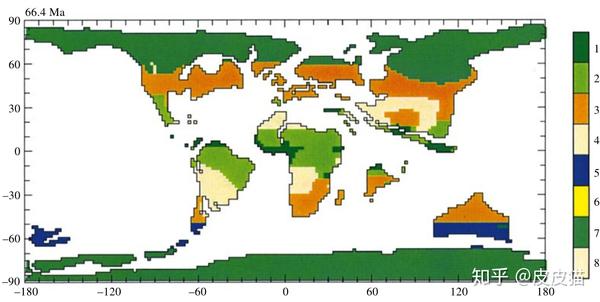
在上图中,北美地区三种主要植被从上往下依次是极地落叶林、亚热带常绿阔叶林和热带半落叶森林。
2.植物种类
自从中侏罗纪被子植物开始出现,到早白垩纪双子叶植物出现,被子植物就开始从热带地区向高纬度入侵,取代原有的裸子和蕨类植物。从阿尔布阶到森诺曼阶,被子植物取代高纬度地区的植被。
毫无疑问,晚白垩纪已是被子植物的天下。虽然被子植物不乏高大的乔木,但大多数仍然是低矮的灌木。森林最高的树木仍然是松、杉等裸子植物。
在种类数量上,北半球晚白垩纪被子植物具有压倒性优势。来自地狱溪组(Hell Creek Formation)的植物化石显示,被子植物的种类占据了90%。
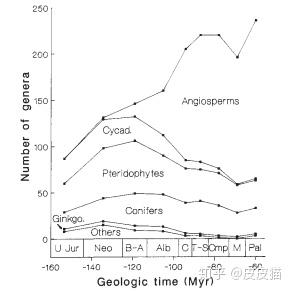
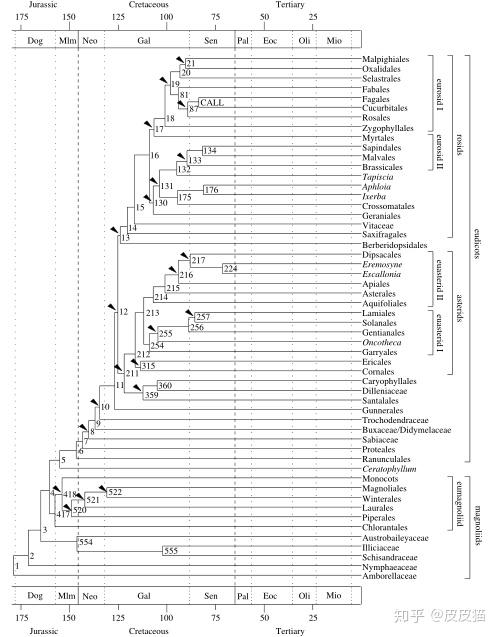
从上图中可以看到,在白垩纪,豆目、葫芦目、蔷薇目、山毛榉目、壳斗目早已出现。这是不是就意味着我们穿越过去仍然可以吃豌豆、板栗、苹果、葫芦、核桃等现代食品?
否。
Dilcher D.(2000)指出,被子植物的第三次共同进化,即多肉的水果和营养丰富的坚果和种子(fleshy fruits and nutritious nuts and seeds)在始新世才完成。

根据Eriksson O.(2000)等人的统计,在马阶晚期,即6600万年前,被子植物的平均果实体积还不到10mm3(立方毫米),跟大米粒的体积差不多。直到始新世,有的果实才能达到我们的拳头那么大。
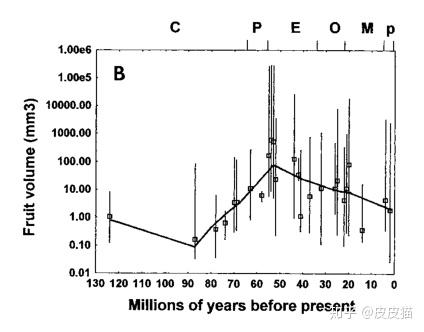
这是由于第三纪鸟类和哺乳动物的崛起,帮助植物散播种子。更大的果实容易被动物吃掉,种子就会被携带到远方。经过长期的协同进化,植物的果实才达到今天的大小。
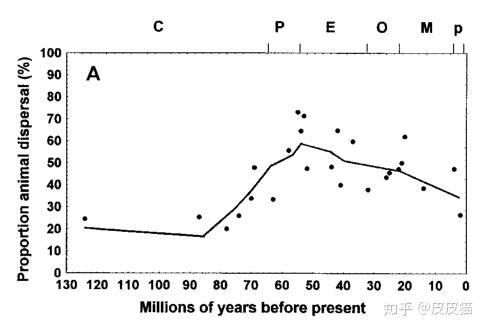
上图与植物果实大型化的曲线是一致的。
既然不能吃水果和坚果,那么磕松子总是可以吧?毕竟裸子植物早就有了。
这个倒是行。
Moles A T(2005)的研究指出,在同时期,裸子植物的种子体积能达到200mm3,差不多就是松子的尺寸。

就是差不多这个东西:

除了磕松子,还能吃银杏。
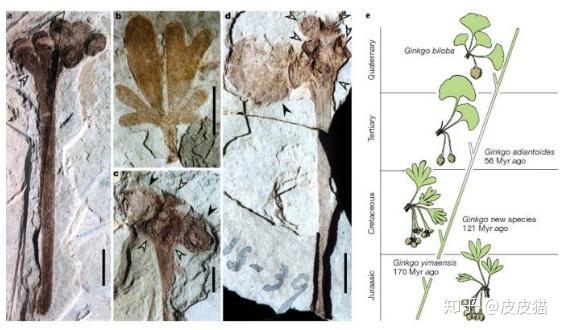
白垩纪银杏在美洲也有分布。种子比今天的要稍小一些,但是仍然可以吃。
由于温暖的气候,中高纬度地区仍然有苏铁。苏铁的种子可以产油,能用来油炸霸王龙腿(想太多)。
总结:
虽然在晚白垩纪末期被子植物已经占据了绝对的优势,但仍然与今天的物种有着很大的区别。最明显的一点就是没有容易食用的种子和果实。许多草本植物可能可以当野菜吃,其块茎可以利用。樟科、八角科植物可以被用作香料(可能)。禾本科植物也已出现,尤其是稻类,不知培养成大米需要多久。这个需要农业专业的人员来解答。
裸子植物与今天差别不大,仍然是松、柏、杉、苏铁、银杏等几种类型,可以像今天一样利用它们。
蕨类植物也有几种,如阴地蕨、链蕨、薇菜、球子蕨、水蕨等几个属,薇菜在今天是可以食用的,水蕨中的满江红也有很大用处,例如饲料。也许穿越者们需要测试这些蕨类的可食用性,然后才能进行培养种植。
————————————————
目录:
皮皮猫:晚白垩纪北美穿越指南——目录————————————————
参考资料:
[1]Wolfe J A, Upchurch Jr G R. North American nonmarine climates and vegetation during the Late Cretaceous[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 1987, 61: 33-77.
[2]Golovneva L B. The Maastrichtian (Late Cretaceous) climate in the northern hemisphere[J]. Geological Society, London, Special Publications, 2000, 181(1): 43-54.
[3]Upchurch G R, Otto-Bliesner B L, Scotese C. Vegetation–atmosphere interactions and their role in global warming during the latest Cretaceous[J]. Philosophical Transactions of the Royal Society of London B: Biological Sciences, 1998, 353(1365): 97-112.
[4]Axelrod D I. Poleward Migration of Early Angiosperm Flora: Angiosperms only displaced the relict Jurassic-type flora at high latitudes in Late Cretaceous time[J]. Science, 1959, 130(3369): 203-207.
[5]Wikström N, Savolainen V, Chase M W. Evolution of the angiosperms: calibrating the family tree[J]. Proceedings of the Royal Society of London B: Biological Sciences, 2001, 268(1482): 2211-2220.
[6]Lidgard S, Crane P R. Quantitative analyses of the early angiosperm radiation[J]. Nature, 1988, 331(6154): 344.
[7]Dilcher D. Toward a new synthesis: major evolutionary trends in the angiosperm fossil record[J]. Proceedings of the National Academy of Sciences, 2000, 97(13): 7030-7036.
[8]Eriksson O, Friis E M, Löfgren P. Seed size, fruit size, and dispersal systems in angiosperms from the Early Cretaceous to the Late Tertiary[J]. The American Naturalist, 2000, 156(1): 47-58.
[9]Chandrasekharam A. Megafossil flora from the Genesee locality, Alberta, Canada[J]. Palaeontographica Abteilung B, 1974: 1-41.
[10]Christophel D C. Fossil floras of the Smoky Tower locality, Alberta, Canada[J]. Palaeontographica Abteilung B, 1976: 1-43.
[11]Moles A T, Ackerly D D, Webb C O, et al. A brief history of seed size[J]. Science, 2005, 307(5709): 576-580.
[12]Prasad V, Strömberg C A E, Leaché A D, et al. Late Cretaceous origin of the rice tribe provides evidence for early diversification in Poaceae[J]. Nature Communications, 2011, 2: 480.
来源:知乎 www.zhihu.com
作者:皮皮猫
【知乎日报】千万用户的选择,做朋友圈里的新鲜事分享大牛。 点击下载
没有评论:
发表评论